FISIOPATOLOGIA
DELLA IPOVISIONE MACULARE
Le
basi fisiopatologiche della riabilitazione visiva
Renato
Meduri
Direttore
della Cattedra di Ottica Fisiopatologica,
Università degli Studi di Bologna
Il danneggiamento di parte dei recetori periferici, di un sistema sensoiale
strutturato, induce innanzitutto una progressiva disattivazione delle reti sinaptiche
encefaliche corrispondenti ai recettori lesi.
Segue un rimodellamento dei circuiti neuronali adiacenti alle aree deprivate.
Si tratta di sequenze neurobiologiche finalizzate a mantenere l’efficacia
informativa dei flussi di stimoli ambientali, sequenze che si inquadrano nel
più ampio contesto della plasticità cerebrale. Questa, di fatto,
alla base dell’apprendimento evolutivo e culturale dell’individuo,
interviene anche in caso di danni subentranti su  sistemi
motori o sensoriali già funzionalmente definiti.
sistemi
motori o sensoriali già funzionalmente definiti.
Acquisizioni elettrofisiologiche, biochimiche, ultrastrutturali, hanno apportato
elementi conoscitivi che permettono già allo stato attuale di inserire
nel bilancio fisiopatogenetico ed evolutivo dei diversi quadri clinici il ruolo,
il livello di intervento, fattori favorenti o inibenti e potenzialità
dei riarrangiamenti sinaptici.
Trattasi di fenomenologie che si attivano anche nel caso di compromessa funzione
retinica maculare e che assumono collocazione nell’ambito clinico e riabilitativo
di tale patologia.
Sequenze sinaptiche visive
I substrati neurali, preposti all’atto percettivo ed alla successiva visione
cosciente, risiedono innanzi tutto nel sistema retinico che recepisce il mondo
esterno in campi recettivi circolari sensibili a forma, colore, movimento.
Quelli per la forma e colore caratterizzano la retina centrale e scaricano sul
sistema ganglionare parvocellulare, mentre quelli per il movimento, più
ampi, occupano la retina paracentrale e periferica e scaricano sul sistema magnocellulare.
I segnali codificati in modulazione di frequenza vengono inviati al corpo genicolato,
che è una struttura subcorticale, mantenendo una organizzazione differenziata:
il sistema magnocellulare afferisce negli strati interni, il parvocellulare
in quelli esterni.
Da qui i segnali giungono alla corteccia striata (V1 o area 17 di Broadman):
gli strati parvocellulari vi acquisiscono sinapsi nello strato IV, il parvocellulare
nel II e III, realizzando una precisa proiezione della retina punto a punto.
Dalla corteccia visiva primaria e ritorno
Sono state identificate specifiche vie percettive che, originando dalla corteccia
visiva primaria, raggiungono aree corticali specializzate per le diverse caratteristiche
dello stimolo.
La via della percezione della forma in movimento ed il movimento, dallo
strato 4 (di V1) di fibre che si portano alle aree parastriate V3 e V5 sia direttamente
sia attraverso relais nell’addensamento di V2 (area corticale che circonda
V1).
La via del colore si proietta sull’area parastriata V4 sia direttam ente
sia dopo connessione con i neuroni delle strisce sottili di V2.
ente
sia dopo connessione con i neuroni delle strisce sottili di V2.
La via della forma associata al colore, deriva da connessioni fra le
regioni intermedie di V1 ed area V4.
In sintesi sono identificabili quattro sistemi specializzati rispettivamente:
per la forma statica, per il colore, per la forma in movimento e per il movimento.
I sistemi per la forma statica e per il colore sono selettivamente parvocellulari,
mentre per la forma dinamica e per il movimento sono selettivamente magnocellulari
e, pur operando in parallelo, hanno caratteristiche funzionali assai diverse.
La velocità di conduzione è lenta nel parvocellulare e rapida
nel magnocellulare; la dinamica di risposta è tonica nel primo e fasica
nel secondo. La sensibilità al contrasto è già satura per
valori del 15% nel magnocellulare mentre nel parvocellulare è oltre l’80%.
La frequenza temporale è alta nel magnocellulare, bassa nel parvocellulare;
la sensibilità cromatica è presente solo nel parvocellulare.
Dalle cortecce specializzate alla corteccia visiva primaria
Alle vie in uscita dalla corteccia V1 corrispondono connessioni di ritorno che,
proiettando dalle aree specializzate, si portano a V1. Tali fibre sono più
diffuse, nel senso che interessano su V1 o a livello di V1 popolazioni
cellulari assai estese ed addirittura su strati diversi (2°, 3°, 4°).
Si realizza così un’integrazione a più stadi in cui il segnale,
dopo una codifica in aree specializzate, viene rinviato, perfezionato, alla
corteccia visiva primaria, integrato ed inserito nella precisa mappa topografica
del mondo esterno caratteristica di V1 (calco retinico). Sotto il profilo funzionale,
percezione e comprensione dell’immagine avvengono contemporaneamente. Va
precisato che i due sistemi magno e parvocellulare funzionano in un fine gioco
di antagonismi con prevalenza fisiologica, di norma, di un effetto inibitore
del secondo sul primo. Nella scala filogenetica, il sistema magnocell ulare
risulterebbe più antico con sviluppo e prevalenza del parvocellulare
solo nelle specie più evolute.
ulare
risulterebbe più antico con sviluppo e prevalenza del parvocellulare
solo nelle specie più evolute.
Di fatto, nell’uomo, la retina non trasmette al cervello il calco fotografico
completo dell’immagine, ma solo taluni punti chiave recepiti dal sistema
parvocellulare, attraverso uno scanning che segue le linee guida dell’immagine
soffermandosi sui particolari significativi. Questi punti, unitamente ai movimenti
di fissazione che l’occhio ha effettuato per raggiungerli, integrati a
livello cerebrale, permettono la ricostruzione delle immagini. Si tratta di
una strategia che realizza l’intervento combinato del sistema parvocellulare
e del sistema magnocellulare e, quindi, una contemporanea percezione sia della
forma che del colore e del movimento.
A livello soggettivo si può avere, tuttavia, l’utilizzo prevalente
di alcune specificità dello stimolo. Un esempio chiarificatore può
derivare dal mondo artistico. Mondrian, nelle sue opere, caratterizzate da viva
cromaticità, fa nettamente prevalere la funzionalità ed il significato
dell’area V4; Tinquely nell’ambito dell’arte cinetica sfrutta,
invece, selettivamente il significato dell’area V3.
Come si giunge alla organizzazione sinaptica
L’assetto sinaptico genicolato e corticale non è genetico, ma deriva
dagli input delle cellule ganglionari retiniche. Già prima della nascita,
quando la retina non ha ancora raggiunto la completa differenziazione degli
strati più esterni, le cellule ganglionari ottiche emettono scariche
regolarmente intervallate secondo uno schema prevedibile e ritmico. Una simile
attivazione inizia il perfezionamento sinaptico della mappa topografica retino-genicolato-corticale.
Dopo la nascita questa attività spontanea è sostitui-ta dallo
stimolo visivo. Si ha così un perfetto adeguamento dell’organizzazione
sinaptica genicolo-corticale alla retina e quindi alle caratteristiche dello
stimolo ambientale.
Tale peculiarità adattativa è determinante quando si consideri
che il flusso di informazioni visive non è né stabile né
costante e che quindi risulterebbe assai difficoltosa, assai dispendiosa, per
l’enorme numero di geni necessari a programmare le opportune connessioni
sinaptiche, una rigida determinazione genetica (l’immagine retinica di
un bambino che nel gioco si allontana e si avvicina presenta una quantità
enorme di variabili e, tuttavia, si continua a classificarlo sempre come bambino
che gioca.
L’induzione fenotipica sinaptica garantisce inoltre la possibilità
di aggiustamenti quando subentrino anomalie del canale informativo. Emblematico
è il cosiddetto riempimento di scotomi del campo visivo: le cellule
retiniche che circondano 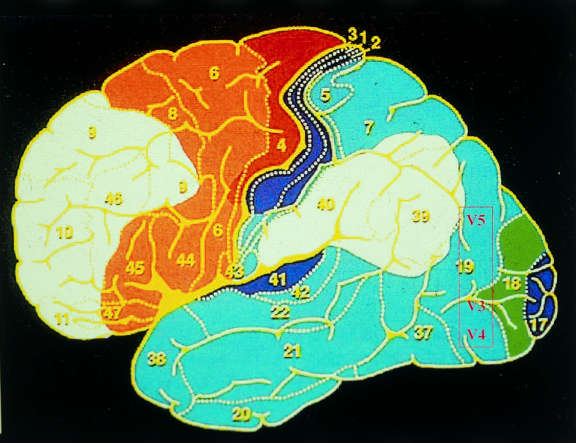 un’area
retinica divenuta insensibile alla luce, molto rapidamente incominciano a connettersi
con i neuroni della corteccia visiva primaria su cui proiettavano le cellule
retiniche lese. E risultato funzionale è una, seppure parziale, neutralizzazione
dell’area scotomatosa.
un’area
retinica divenuta insensibile alla luce, molto rapidamente incominciano a connettersi
con i neuroni della corteccia visiva primaria su cui proiettavano le cellule
retiniche lese. E risultato funzionale è una, seppure parziale, neutralizzazione
dell’area scotomatosa.
Tale plasticità, che si attua in soggetti adulti con strutture neuronali
già da tempo operanti, secondo schemi indotti durante la fase di sviluppo,
è comprensibile considerando i meccanismi attraverso cui le giunzioni
sinaptiche si definiscono per effetto dello stimolo. Si tratta di aggiustamenti
molecolari e neuronali che vedono come protagoniste primarie le proteine costituenti
i ricettori NMDA (N-Metil-Di-Aspartato) dei canali della membrana neuronale
postsinaptica che controllano l’ingresso degli ioni di calcio. Questi recettori
per attivarsi necessitano di due input: una depolarizzazione della membrana
ed in concomitanza un legame con il glutammato.
Questi recettori funzionano così come veri e propri interruttori di coincidenze
idonei alla stabilizzazione sinaptica. In termini informatici le sinapsi nervose
possono venire rappresentate come entità in situazione funzionale binaria,
rispettivamente: attività o attività potenziata. Di fatto in seguito
ad uno stimolo ripetuto l’efficienza sinaptica progressivamente aumenta,
potenziandosi, potendo raggiungere un incremento di superficie di due volte
e mezza. Il fenomeno è noto come fissurazione. Da tale situazione, se
permane lo stato di stimolo, si ha una vera e propria gemmazione sinaptica con
possibile connessione dei neuroni in un nuovo circuito locale. Tale peculiarità
si mantiene anche in età avanzata. Quando, invece, uno dei due segnali,
nel caso di turbe del recettore periferico (la depolarizzazione), venga a decadere,
questa organizzazione si attenua e si rendono possibili nuove organizzazioni
per stimoli coincidenti. Questo può essere accelerato ed ottimizzato
con input adeguati e reiterati.
I compensi in caso di lesioni maculari
In caso di lesioni maculari estese, che compromettono quindi gravemente il sistema
parvocellulare e la corrispondente attività delle aree specializzate
parastriate (V4) per la forma statica ed il colore si ha un prevalere compensatorio
delle attività magnocellulari deputate alla forma dinamica. È
osservazione comune che i pazienti con degenerazione maculare utilizzino l’attività
di V3 deputata alla percezione della forma in movimento: vedono agevolmente
i volti in movimento, come quello delle immagini televisive, mentre dimostrano
incapacità ad individuare i volti di amici e parenti, ricorrendo sovente
a movimenti del capo per facilitarne la comunicazione.
Un ulteriore riarrangiamento è lo svilupparsi di un punto di fissazione
preferenziale, che presuppone, di fatto, una vera e propria rimappatura proiezionale
retino-corticale.
Fattori neuroendocrini
Componenti neuroendocrini possono esplicare effetto positivo o negativo sulle
dinamiche organizzative sinaptiche.
Un ruolo peculiare è da attribuire ai complessi sistemi di innervazione,
che da neuroni sottocorticali (tronco cerebrale), controllano la cortecc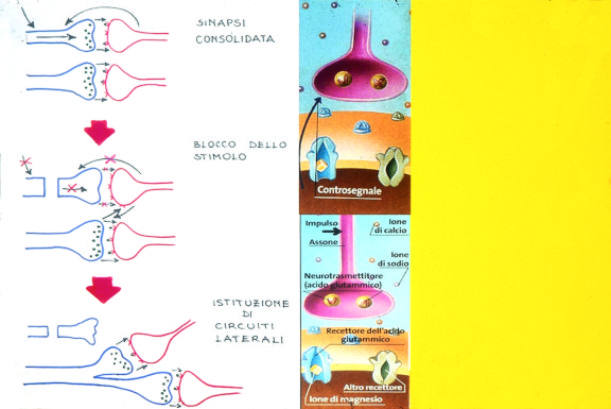 ia
con liberazione di neurotrasmettitori. In relazione al tipo di neurotrasmettitore
sono individuabili cinque principali sistemi sottocorticali, in relazione al
tipo di neurotrasmettitori che gestiscono: il noradrenergico, il serocolinergico,
il dopaminergico, il colinergico, il gabaergico. Oltre a questi sistemi, che
modulano ampie e generiche funzioni cerebrali, vi sono altri sistemi, il cui
numero è in aumento con attività di controllo più selettivo.
ia
con liberazione di neurotrasmettitori. In relazione al tipo di neurotrasmettitore
sono individuabili cinque principali sistemi sottocorticali, in relazione al
tipo di neurotrasmettitori che gestiscono: il noradrenergico, il serocolinergico,
il dopaminergico, il colinergico, il gabaergico. Oltre a questi sistemi, che
modulano ampie e generiche funzioni cerebrali, vi sono altri sistemi, il cui
numero è in aumento con attività di controllo più selettivo.
È opportuno considerare tali aspetti in quanto questi sistemi sottocorticali
subiscono, in caso di handicap visivo, scompensi funzionali che possono rendere
difficoltosa, e comunque non soddisfacente, l’efficacia dell’iter
riabilitativo. Almeno per quanto concerne i sistemi principali sussiste la possibilità
di vagliarne l’equilibrio con dosaggi endocrini degli ormoni ipofisari
e corticosurrenali. Nel caso di squilibri la moderna farmacologia permette soddisfacenti
compensi.